Om felles avstamning -debattinnlegg:
Svar til et innlegg fra Hans Øvland Tjelle.
1. Morfologi: Homologi (strukturell likhet) er likhet i hvordan kroppsdeler er bygd opp. Denne likhet tar neo-darwinismen til inntekt for at organismene er beslektet- et alternativ er felles design. Fossilmaterialet er forenlig med begge forklaringer. Neo-darwinismen tar homologi som et bevis for felles avstamning. En legger inn i premisset at homologi skyldes felles opphav. Når en så finner tegn på homologi, tolker en det som felles avstamning. Det blir sirkelargumentasjon der en går ut fra det en skal bevise. En enkel oppstilling av former som ligner hverandre, gir ikke en forklaring på opphavet i seg selv. Det kreves en mekanisme som kan forklare slike likheter.
En har antatt at DNA-likheter er bedre argument enn strukturell (morfologisk) likhet; Spesielt gjelder det siden analoge strukturer, som virker likeformet -eksisterer, uten at noen mener det skyldes felles avstamning. Men heller ikke det har ført fram. En har funnet at: Homologe gener ikke nødvendigvis koder for homologe strukturer. Det er heller regel enn unntak at homologe strukturer stammer fra klart ulike utgangspunkt. Genet ‘sonic hedgehog’ er ett unntak i så måte. Hoved-konklusjon: homologi kan ikke brukes som bevis for sannheten i evolusjonslæren.
Bilde 1. Eks. på 'konvergent evolusjon'
2. Genetikk - Cytochrom C: Her er et problem for evolusjonistene hvordan 'molekylklokken' kunne ha krysset så jevnt i et gitt protein i så mange forskjellige organismer (til tross for noen anomalier som tidligere ble diskutert som gir enda flere problemer). For at dette skal fungere, må det være en konstant mutasjonsrate per enhetstid over de fleste typer organismer. Men observasjoner viser at det er en konstant mutasjonsrate per generasjon, så det burde være mye raskere for organismer med rask generasjonstid, for eksempel bakterier, og mye langsommere for elefanter. I insekter varierer generasjonstider fra uker i fluer i mange år i cikader, og likevel er det ingen bevis for at fluer er mer divergerte enn cikader. Så bevisene er mot teorien om at de observerte mønstrene skyldes mutasjoner som akkumuleres over tid etter hvert som livet utviklet seg.
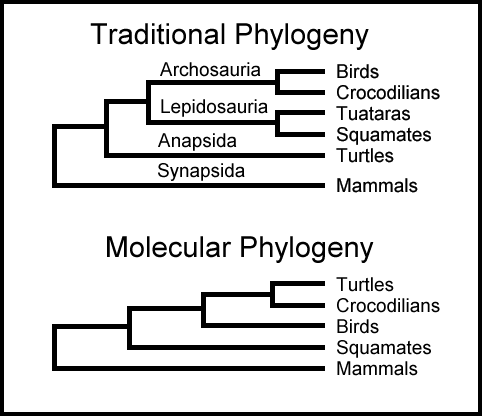 3. Fylogenetiske trær: DNA-data leder ofte til konfliktfylte fylogenetiske kronologier. F.eks. viser Collins evolusjonære tre i "The language of God", at flyvende lemurer slekter på ekornlignende dyr, og kaniner og aper til mer fjerne grener. Men i en fylogenetisk studie publisert i 2002 (Proceedings of the National Acedemy of Sciences USA) viser at flyvende lemurer slekter mer på aper og de ekornlignende dyrene (tree shrews) mer er i slekt med kaniner. Dessuten er det ofte konflikt mellom fylogeneser basert på DNA i forhold til anatomiske baserte fylogeneser. For eks. for hvaler, som i 1960-årene ble påstått å stamme fra Mesonychider, en utdødd gruppe kjøttetende hovdyr. Men i 1990-årene viste molekylære fylogeneser større likhet til flodhest. I 2007 pekte J.G.M Thewissen og kolleger på at siden hvaler dukker opp i fossil-materialet 35 millioner år tidligere enn flodhesten, er det ikke sannsynlig at de to er nær relatert. Det fremgår betydelig mer i 2007 analysen fra britiske forskere enn den omforente konklusjonen:
3. Fylogenetiske trær: DNA-data leder ofte til konfliktfylte fylogenetiske kronologier. F.eks. viser Collins evolusjonære tre i "The language of God", at flyvende lemurer slekter på ekornlignende dyr, og kaniner og aper til mer fjerne grener. Men i en fylogenetisk studie publisert i 2002 (Proceedings of the National Acedemy of Sciences USA) viser at flyvende lemurer slekter mer på aper og de ekornlignende dyrene (tree shrews) mer er i slekt med kaniner. Dessuten er det ofte konflikt mellom fylogeneser basert på DNA i forhold til anatomiske baserte fylogeneser. For eks. for hvaler, som i 1960-årene ble påstått å stamme fra Mesonychider, en utdødd gruppe kjøttetende hovdyr. Men i 1990-årene viste molekylære fylogeneser større likhet til flodhest. I 2007 pekte J.G.M Thewissen og kolleger på at siden hvaler dukker opp i fossil-materialet 35 millioner år tidligere enn flodhesten, er det ikke sannsynlig at de to er nær relatert. Det fremgår betydelig mer i 2007 analysen fra britiske forskere enn den omforente konklusjonen:
Se mer om det her.
Bilde 2. Ulike metoder -ulike resultat
Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Woese tok hovedsakelig fatt på de uoverensstemmelser som befinner seg på systematikkens høyere nivåer, 'rikene'. Men problemet berører også mindre grener, herunder fylogenese for dyr. Biologen Michael Lynch skrev i 1999: "Å få klarhet i de større dyrerekkenes innbyrdes forbindelser har vist seg være et problem det er vanskelig å definere nærmere". Analyser av forskjellige gener, ja selv analyser av de samme gener har bragt en mengde ulike fylogenetiske stamtrær for dagen. Selv i de tilfeller der ulike molekylanalyser kan settes sammen, er resultatet ofte bisart.
F.eks. plasserer en studie fra 1996 som har anvendt 88 proteinsekvenser, kaniner i samme gruppe som primatene i stedet for hos gnagerne. En analyser fra 1998 plasserer kua i nærmere slektskap med hvalen enn med hesten. Den molekylære fylogenesen synes være blitt plassert i en sump med mudder til halsen.
-------
 Om 'urfuglen' (Archaeopteryx) og dens status som mellomform mellom krypdyr og fugler. Den var uten tvil en fugl: den hadde fjær og ikke skjell. Den hadde nebb og ingen kjeve. Den var ganske sikkert varmblodig, og ikke vekselvarm. Den hadde vinger med fjær, som den flakset med når den fløy. Intet krypdyr har hatt fuglevinger. Flyveøgler hadde svevehud, i likhet med flaggermus. Det finnes likefullt felles trekk med fugler og dinosaurer: Archaeoteryx har en ikonisk status i darwinismen. Den betraktes som et 'endegyldig bevis' på overgangen mellom krypdyr og fugler. Den visker liksom ut alle grenser og gjør overgangen mellom krypdyr og fugler til en 'naturlig og hverdagslig' sak.
Om 'urfuglen' (Archaeopteryx) og dens status som mellomform mellom krypdyr og fugler. Den var uten tvil en fugl: den hadde fjær og ikke skjell. Den hadde nebb og ingen kjeve. Den var ganske sikkert varmblodig, og ikke vekselvarm. Den hadde vinger med fjær, som den flakset med når den fløy. Intet krypdyr har hatt fuglevinger. Flyveøgler hadde svevehud, i likhet med flaggermus. Det finnes likefullt felles trekk med fugler og dinosaurer: Archaeoteryx har en ikonisk status i darwinismen. Den betraktes som et 'endegyldig bevis' på overgangen mellom krypdyr og fugler. Den visker liksom ut alle grenser og gjør overgangen mellom krypdyr og fugler til en 'naturlig og hverdagslig' sak.
Bilde 3. Fuglevinger og nebb = fugl
Museumsbestyrer Colin Patterson som i sin tid oppbevarte Archaeopteryx: "Jeg vil ikke stikke noe under stolen: Det finnes ikke her ett eneste sådant fossil (som brukes som 'stamfar'eller overgangsform), hvormed man vil kunne lage et vanntett argument". Det er interessant i og med den såkalte 'urfuglen' befant seg i han varetekt på Natual History Museum. Her skal også tas med Patersons begrunnelse, som jeg nå er gjort oppmerksom på: "“Årsaken er at utsagn om forfedre og nedstigning ikke er aktuelt i fossilregistreringen. Er Archeopteryx forfedre for alle fugler? Kanskje ja, kanskje nei: det er ingen måte å svare på spørsmålet på." (1) {Dette skyldes bl.a. at fossiler ikke kommer med ID-papirer, og det ikke er mulig i utgangspunktet og si at det ene fossilet stammer direkte fra et annet -oversetters kommentar.}
I og med at Archaeopteryx ofte nevnes som en overgangsform mellom krypdyr og fugler, er det viktig å skjelne begrepsmessig mellom mellomliggende former og overgangsformer. En mellomliggende form er hva ordet sier, en form som vil kunne plasseres mellom to av disse kriterier, A og B. Det behøver derimot ikke indikere at det kan påvises at den stammer fra A og er opphav til B. Hvis en hevder noe slikt, trenger en også kunne påvise en mekanisme som er egnet til denne oppgaven. Heller enn en overgangsart mellom krypdyr og fugler, holder de fleste paleontologer nå for at Archaeopteryx er en unik mellomform, en art av fugler som nå er utdødd.
Om at 'fugler er etterkommere av dinosaurer':
En artikkel i Nature -her, "A new hypothesis of dinosaur relationships and early dinosaur evolution," presenterer nye ideer om dinosaur-relasjoner som avslører i hvilken grad dinosaur egenskaper ikke er fordelt i et trelignende mønster. En nyhetsartikkel kaller dette en "radikal rystelse av dinosaur familietreet" fordi det ville snu om på et århundre med evolusjonær tenkning omkring dinosaurer:
Analysen, som allerede har skapt kontrovers i den akademiske verden, tyder på at de to grunnleggende grupper som dinosaurer er klassifisert på i mer enn et århundre, trenger en grunnleggende revurdering. Hvis den viser seg riktig, vil den reviderte versjonen av slektstreet styrte noen av de mest grunnleggende antakelser om dette kapitlet av evolusjonær historie, inkludert hvordan felles stamfar til alle dinosaurer så ut og hvor den kom fra. ..
Bilde 4. Skulle fugler være i dennes 'avstamningslinje'?(sinosauropteryx-prima)

Vurder disse kommentarer av studiens hovedforfatter, Cambridge University graduate student Matthew Baron: "Funnene støtter også den mulighet at dinosaurer som Stegosaurus og Triceratops, tradisjonelt fremstilt som tanklignende pansrede dyr, kan ha hatt fjær" .. "Kanskje hadde vi dunkledde Triceratops og Stegosaurus," sa Baron. "Det kan være at fjærene ville blitt stukket ut mellom skjellene, kunne det ha vært en vakker dunkledd fargerik fjærdrakt ... eller skjell dekket i dunete fjær. Det er mulig." Slik grunnløse spekulasjoner antyder hvilke agendaer som virkelig kan drive den nye klassifiserings ordningen.
Mer om dette her.
Til slutt noe om hvorvidt ID møter krav om testbarhet og falsifisering:
 Endring fra en kroppsplan, for eksempel en svamp eller en orm, til en annen kroppsplan, slik som fisk, krever mange dramatiske endringer for å bli gjennomført, på en gang, gjennom intelligent veiledning. Denne konklusjonen fører direkte til en forventning om at nye kropps- planer hos phyla (dyrerekker) plutselig skal vises i fossilregisteret, uten en sammenhengende serie av mellomprodukter, som fører tilbake til stammen av et evolusjonært tre. Og dette er hva vi finner, se f.eks. her og her..
Prediksjonen passer perfekt mønsteret sett i den kambriske eksplosjonen -her, og i senere plutselige opptredener av nye arkitekturer. Mens det lenge var et evolusjonistisk glansnummer ('evolusjon i hullene') at junk-DNA utgjorde storparten, mens en i følge ID forventet at det aller meste ville ha funksjoner. Denne forutsigelsen er nå bekreftet.
Endring fra en kroppsplan, for eksempel en svamp eller en orm, til en annen kroppsplan, slik som fisk, krever mange dramatiske endringer for å bli gjennomført, på en gang, gjennom intelligent veiledning. Denne konklusjonen fører direkte til en forventning om at nye kropps- planer hos phyla (dyrerekker) plutselig skal vises i fossilregisteret, uten en sammenhengende serie av mellomprodukter, som fører tilbake til stammen av et evolusjonært tre. Og dette er hva vi finner, se f.eks. her og her..
Prediksjonen passer perfekt mønsteret sett i den kambriske eksplosjonen -her, og i senere plutselige opptredener av nye arkitekturer. Mens det lenge var et evolusjonistisk glansnummer ('evolusjon i hullene') at junk-DNA utgjorde storparten, mens en i følge ID forventet at det aller meste ville ha funksjoner. Denne forutsigelsen er nå bekreftet.
Bilde 5. Noen mener ID falsifiseres ved slike bilder (ubeskrevet fossil)
Mer om hvorvidt ID har kan falsifiseres, kan en lese her og evt. her.
Utvalg og bilder ved Asbjørn E. Lund